Laboratoř buněčné biologie
Přehled laboratoře
(další informace naleznete také na našich webových stránkách www.cell-biology.cz)
Naším hlavním úkolem je identifikovat a pochopit molekulární moduly, které regulují polaritu a morfogenezi rostlinných buněk, se zaměřením na ty, které fungují na křižovatce sekreční dráhy, lipidů plazmatické membrány a aktinového cytoskeletu. Naši pozornost rozdělujeme mezi dva hlavní procesy, které dávají rostlinným buňkám jejich tvar: orientované buněčné dělení a diferencovaný buněčný růst, a zaměřujeme se na nitrobuněčné molekulární mechanismy řídící buněčnou morfogenezi, jako je exocytóza.

Náš výzkum
Organizace buněčných procesů u eukaryot závisí na transportu extracelulární matrix, membránových lipidů a proteinů na povrch buňky, který se uskutečňuje převážně prostřednictvím exocytotických vezikul. Vrcholem sekreční dráhy je exocytóza, jemně vyladěný proces zahrnující přivázání, připojení a splynutí sekrečních vezikul s plazmatickou membránou, který nakonec vyvrcholí uvolněním nákladu.
Jádrem našeho zkoumání je exocyst, konzervovaný oktamerický proteinový komplex, který hraje ústřední roli při připoutávání sekrečních váčků k plazmatické membráně, a slouží tak jako klíčový regulátor exocytózy. Naše výzkumné úsilí se soustřeďuje na rozbor funkčních aspektů role exocystu u různorodého spektra rostlinných druhů, od krytosemených rostlin Arabidopsis thaliana a Nicotiana tabacum až po mechorosty Physcomitrium patens a Marchantia polymorpha, a streptofytní řasu Klebsormidium nitens. Důležitým aspektem našeho výzkumu je zkoumání úlohy vezikulárního transportu, konkrétně exocystu, v reakci rostlin na abiotické stresy a v interakcích mezi rostlinami a mikroorganismy.
Významné výsledky 2021-2023
Molekulární vhled do funkce a evoluce komplexu exocyst u rostlin
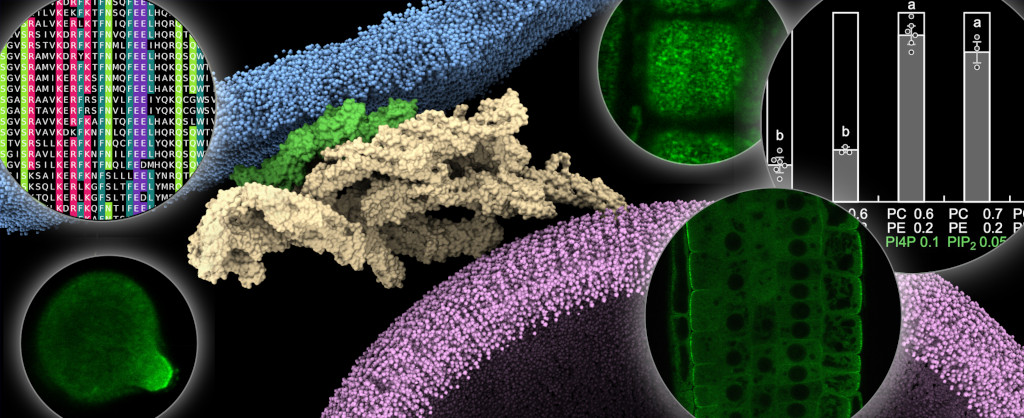
Obrázek 1: Model naznačující, jak komplex exocyst u Arabidopsis připojuje sekreční váček k plazmatické membráně prostřednictvím izoformy EXO70A1 a několika záporně nabitých fosfolipidů. Vložené obrázky ukazují mnohostranný metodický přístup naší laboratoře, který kombinuje nejmodernější mikroskopické, výpočetní, biochemické a in planta techniky. Inspirováno výsledky publikovanými v (Synek, Pleskot, Sekereš et al. 2021).
Objasnili jsme evoluční historii komplexu exocyst suchozemských rostlin a odhalili vícestupňový vzorec evoluce jeho různých podjednotek. Rozčlenili jsme evoluci podjednotky EXO70 a odhalili její rozdělení na tři dobře zavedené podrodiny, z nichž každá vykazuje jedinečné funkční vlastnosti. Pomocí mezidruhových komplementačních analýz jsme poukázali na hlubokou konzervaci kanonické podrodiny EXO70.1 a nezávislou evoluci nekanonických podrodin EXO70.2 a EXO70.3 (Haluška, Janková-Drdová et al., submitted). Kromě toho naše zkoumání izoforem podjednotek exocystu SEC15a a SEC15b osvětlilo jejich hlavní role v samčím gametofytu a sporofytu, a odhalilo překvapivé získání nové funkce pro SEC15a ve sporofytu (Batystová et al. 2022). Naše funkční studie exocystu u mechu Physcomitrium patens přinesla poznatky o významu podjednotky exocystu SEC6 v buněčném dělení a organizaci mnohobuněčných struktur. Naše pozorování dokumentují roli komplexu exocyst při přechodu od jednoduchých vláknitých struktur ke složitému morfologickému uspořádání v rostlinných orgánech (Obr. 2) (Brejšková et al. 2021).

Obrázek 2: Narušení komplexu exocyst u mechu Physcomitroium patens vede k závažným vývojovým vadám. Převzato z (Brejšková et al. 2021).
Úloha vezikulárního tranportu v interakcích mezi rostlinami a mikroby
V reakci na patogenní houby využívají rostliny intenzivní exocytózu k ukládání výztuh buněčné stěny (papil nebo tzv. encasementů), které zamezují šíření infekce po celé rostlině. Identifikovali jsme zásadní roli komplexu exocyst obsahujícího podjednotku EXO70B2 v membránových doménách papil důležitých pro ukládání kalózy a tvorbu inkrustací buněčné stěny. Naše výsledky poukázaly na význam exocystu obsahujícího EXO70B2 při doručování klíčového proteinu SNARE SYP121 do membránových domén papil, což nabízí nové poznatky o mechanismech odolnosti vůči pronikání. (Ortmannová et al. 2022). Zjistili jsme, že blízce příbuzná podjednotka exocystu EXO70B1 hraje zásadní roli v reakcích na různé abiotické stresy, přičemž působí jak v endomembránách, tak na plazmatické membráně ve složitém režimu vzájemné spolupráce/kompetence s podjednotkou EXO70B2 (Drs et al. v přípravě).
Náš výzkum objasnil buněčný procesing, transport, lokalizaci, sekreci a funkci proteinu PR1 (pathogenesis-related 1). Vytvořením spektra značených variant PR1 v Arabidopsis jsme nastínili vhled do dynamické modulace obranných reakcí a imunitních drah prostřednictvím zpracování proteinu PR1. Tato složitá souhra je závislá na buněčné lokalizaci a stáří rostliny, což zdůrazňuje mnohostrannou povahu tohoto klíčového obranného proteinu rostlin (Pečenková et al. 2022).
Abychom odhalili vzájemný vztah mezi morfogenezí buněk a reakcemi na biotický stres, začali jsme zkoumat, jak elicitor imunity chitosan ovlivňuje polární růst rostlinných buněk. Zjistili jsme, že chitosan vyvolává ukládání kalózy v kořenových vláskách a inhibici růstu. Naše zjištění odhalila hluboké zachování této strategie v analogických strukturách podobných kořenovým vláskům u lýkožroutů a mechorostů, což podtrhuje význam této reakce v situacích mírného biotického stresu. (Drs et al., submitted).
Lipidová signalizace na plazmatické membráně rostlin
Růst pylové láčky, klíčové součásti pohlavního rozmnožování rotlsin, vyžaduje přísně regulovaný mechanismus vylučování pektinu k udržení plasticity buněčné stěny. Na této regulaci se na apikální plazmatické membráně podílejí fosfoinositidy a kyselina fosfatidová, ale procesy regulující jejich produkci zůstávají nejasné. Ve spolupráci se skupinou Tilla Ischebecka z univerzity v německém Göttingenu jsme ukázali, že diacylglycerolkináza 5 hraje významnou roli v regulaci sekrece pektinu v apikálně rostoucích pylových láčkách (obr. 3) (Scholz, Pejchar et al. 2022).
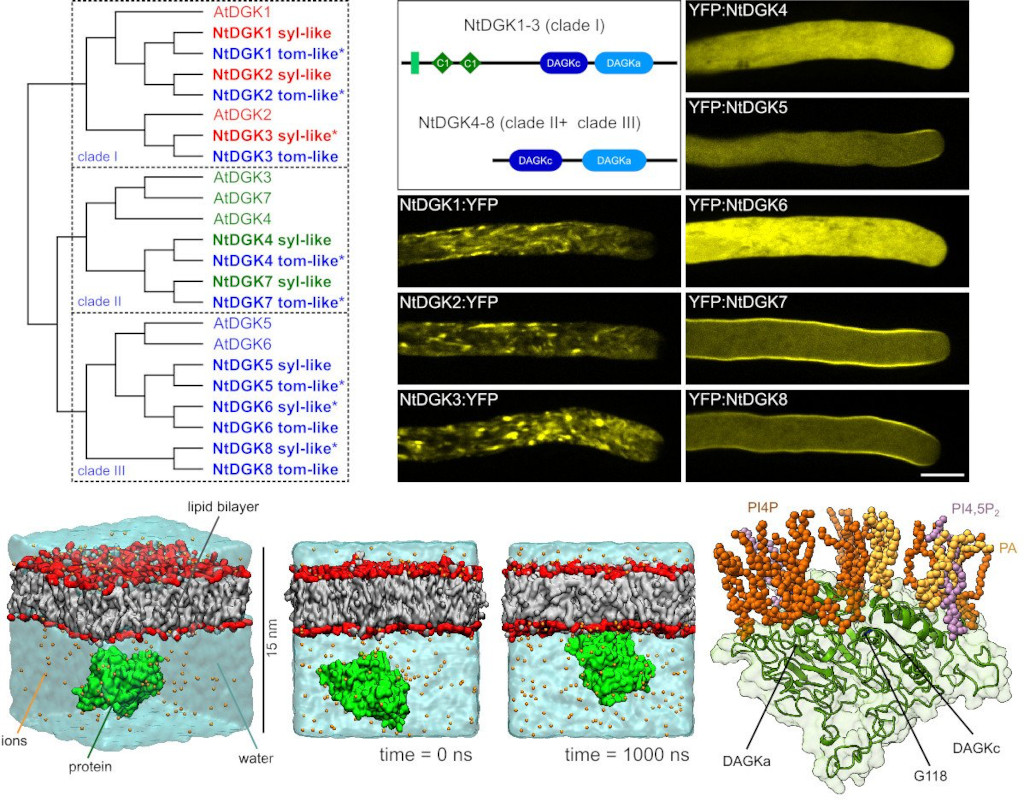
Obrázek 3: Izoformy tabákové diacylglycerol kinázy vykazují odlišné lokalizační vzorce v pylových trubicích a vážou se na plazmatickou membránu prostřednictvím fosfoinositidů. Vlevo fylogeneze izoforem diacylglycerol kinázy (DGK) Arabidopsis a Nicotiana tabacum a jejich rozdělení do tří kladů. Vpravo rozložení domén a lokalizace NtDGK1-8 značené YFP v aktivně rostoucích pylových láčkách. Dolní panel znázorňuje simulace molekulární dynamiky ukazující mechanistické detaily interakce NtDGK5 s membránou. Měřítko 10 μm. Převzato z (Scholz, Pejchar et al. 2022).
Protože membránové lipidy jsou důležitými hráči v reakcích rostlin na různé stresy, zkoumali jsme zapojení lipidové signalizace do vnímání signalizace flagelinu, jednoho ze základním kamenů rostlinné imunity vyvolané takzavnými molekulárními vzory (MAMPs). Ve spolupráci s Laboratoří patologické fyziologie rostlin a skupinou Erica Ruellanda (CNRS, Compiègne, Francie) jsme zjistili, že peptid flg22 odvozený od flagelinu způsobuje u rostlin Arabidopsis rychlé a přechodné změny v dynamice lipidů. Jako gen odpovědný za tyto změny jsme identifikovali diacylglycerol kinázu 5 (DGK5), enzym produkující kyselinu fosfatidovou. Mutantní rostliny dgk5.1 s porušenou funkcí DGK5 produkovaly v reakci na flg22 méně kyseliny fosfatidové a vykazovaly zhoršenou odolnost. Enzymatická aktivita DGK5 lokalizovaného v plazmatické membráně je tedy zásadní pro signalizaci flagellinu a časné imunitní odpovědi při interakcích mezi rostlinami a mikroby (Kalachova, Škrabálková et al., 2022).

Obrázek 4: Ukázky titulních stran odborných časiopisů publikovaných v letech 2021-2203, které upozorňují na výsledky naší práce (Batystová et al. 2022, Synek et al. 2021, Markovic et al. 2020).
Pracovníci
vedoucí skupinyvedoucí vědecký pracovník
- Ing. Martin Potocký Ph.D.
zástupce vedoucího laboratořevědecký pracovník
- Tamara Pečenková Ph.D.
vědecký pracovník
- Mgr. Edita Drdová
- RNDr. Michal Hála Ph.D.
- Ing. Přemysl Pejchar Ph.D.
- Mgr. Lukáš Synek PhD.
- RNDr. Viktor Žárský CSc.
vědecký asistent
- Mgr. Lucie Brejšková PhD.
- Ing. Andrea Potocká Ph.D.
- Mgr. Hana Soukupová Ph.D.
postdoktorand
- Mgr. Jitka Ortmannová PhD.
- MSc. Ankush Saddhe PhD
doktorand
- Mgr. Klára Batystová
- Msc. Laiju Chandran Ka...
- Bc Matěj Drs
- Samuel Haluška
- Ing. Ondřej Novotný
- Mgr. Eliška Škrabálková
laborant
- Bc. Jana Šťovíčková
Výzkumné projekty
- All roads lead to ROS: spatio-temporal regulation of pollen NADPH oxidases (Czech Science Foundation - GAČR project GA , GA ČR , Přemysl Pejchar
- FASS/TON2 functions in moss cell morphogenesis and phylogenetic insights into the TTP complex evolution , GA ČR , Viktor Žárský
- The plasticity of protein-lipid interfaces in plant membranes perceived through evolutionary lenses , GA ČR , Martin Potocký
- Good-Cop/Bad-Cop: Distinct roles of anionic phospholipids in plant endocytosis , GA ČR , Martin Potocký
- SIGNALS - The role of anionic phospholipids in the regulation of endocytosis in tip-growing plant cells , MŠMT , Martin Potocký
- Control of plant exocyst function by protein phosphorylation in root hairs and pollen tubes - role of unconventional exocyst complex subunits EXO70C , GA ČR , Viktor Žárský
- Multifaceted analysis of diacylglycerol kinase family in plants , GA ČR , Přemysl Pejchar
- Plant exocyst complex function in autophagy-related membrane transport , GA ČR , Tamara Pečenková
- Multiscale analysis of signalling phospholipids and their interaction protein partners in the regulation of plant tip growth , GA ČR , Martin Potocký
- Funkce rostlinného poutacího komplexu exocyst v exocytóze, buněčném dělení a biogenezi buněčné stěny , GA ČR , Michal Hála
- Úloha diacylglycerolu při toxickém působení hliníku u rostlin , GA ČR , Přemysl Pejchar
- Exocyst, poutací komplex sekretorických váčků, v polarizaci transportu auxinu , GA ČR , Lukáš Synek
- Charakterizace vybraných příslušníků nových skupin rostlinných forminů - třídy II a třídy III. , GA ČR , Viktor Žárský
- Chracterisation of NADPH oxidase from tobacco pollen and its role in regulation of polar cell expansion , GA ČR , Martin Potocký
- Transkriptom mutanta exo70A1 a buněčné funkce EXO70A1, předpokládané podjednotky exocystu , GA AV , Lukáš Synek
- Transkriptom mutanta exo70A1 a buněčné funkce EXO70A1, podjednotky komplexu exocyst, u Arabidopsis thaliana , GA AV , Lukáš Synek
Publikace
2014
- FRONTIERS IN PLANT SCIENCE 5 5 2014 (External fulltext)
- Plant Cell Morphogenesis: Methods and Protocols 1080 231-236 2014 (External fulltext)
- NEW PHYTOLOGIST 203 483-494 2014 (External fulltext)
- ACTA PHYSIOLOGIAE PLANTARUM 36 1981-1991 2014
- DEVELOPMENTAL CELL 29 607-620 2014 Fulltext: veřejný PDF odkaz
- FRONTIERS IN PLANT SCIENCE 4 543 2014
- 2014
2013
- PROGRESS IN LIPID RESEARCH 52 62-79 2013 (External fulltext)
- TRENDS IN PLANT SCIENCE 2013 (External fulltext)
- In: Plant Lipid Signaling Protocols eds. Munnik T. Heilmann I. (Humana Press New York) 193-203 2013 (External fulltext)