Aminokyselina jako přepínač citlivosti
Brefeldin A (BFA) je syntetizován mnoha houbovitými organizmy. Původně byl izolován a charakterizován jako antibiotikum, antivirotikum a jako fungicid. Záhy však bylo objeveno, že umí blokovat sekreci bílkovin. BFA zabraňuje exportu bílkovin z buněk, především působením na Golgiho aparát a způsobuje přerozdělení bílkovin z Golgiho aparátu do endoplazmatického retikula. V Laboratoři hormonálních regulací se BFA používá při studiu transportu bílkovin a váčků v buňkách.
Citlivost vůči BFA je v buňkách závislá na jedné malé bílkovině, obecně nazývané ARF-GEF. Tato bílkovina zajišťuje výměnu „energii nesoucích“ molekul GTP za GDP na větších bílkovinách, tzv. ARFech. Když se tato výměna uskuteční, mohou ARFy konat svou práci. Například umožní vychlípit membránu, aby se následně mohl vytvořit váček nesoucí rozličné bílkoviny do různých míst v buňce (tedy na různé adresy). Nebo zajistí přišpendlení určité adresy k váčku, aby se váček ve spleti transportních drah v buňce nezatoulal. Je známo, že když bílkovina ARF-GEF má v místě, které je důležité pro výměnu GTP za GDP, aminokyselinu leucin, pak se BFA na toto místo váže a blokuje výměnu a následná dráha je blokována. Jestliže je na tom samém místě aminokyselina metionin, pak je ARF-GEF k BFA necitlivý a ARF koná práci, jako by BFA nebyl přítomný. Například u modelového huseníčku blokuje BFA transport váčků ven z buňky. Endocytóza (tvoření váčků na plasmatické membráně a jejich putování dovnitř do buněk, tudíž opačný směr) zůstává nenarušena.
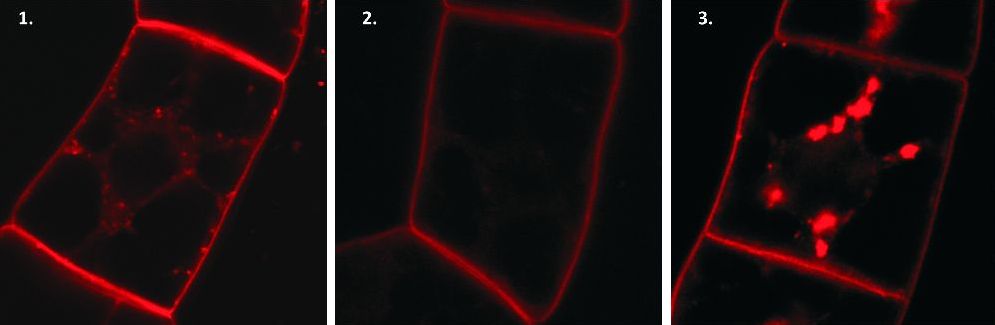
Zjistili jsme, že po přidání BFA k našim oblíbeným buňkám tabáku se přeruší také tok váčků od plazmatické membrány směrem do buňky (obr. 1) a červená barva, která normálně ve váčcích putuje do buňky, zůstává zadržena na membráně (obr. 2). Jinými slovy BFA tedy u buněk tabáku inhibuje i endocytózu. Podívali jsme se do databáze ARF-GEFů tabáku a zjistili jsme, že všech osm nalezených má ve vazebném místě leucin. Nahradili jsme tedy u jedné z vybraných ARF-GEF bílkovin leucin metioninem a „vložili“ ji do buněk tabáku. Díky této výměně se ARF-GEF stal necitlivým k přítomnosti BFA. A opravdu, váčky se u takto transformovaných buněk tvořily na plazmatické membráně a putovaly dovnitř buňky (obr. 3). Díky ostatním, BFA citlivým ARF-GEFům se tyto váčky následně začaly hromadit okolo buněčného jádra, protože BFA zablokoval jejich funkci. Abychom tomuto zabránili, mohli bychom vybrat zas jiný BFA-citlivý ARF-GEF tabáku, „znecitlivět“ ho vůči BFA a umožnit mu tak konat svou práci i v přítomnosti BFA, třeba podílet se na putování váčků směrem od jádra k plasmatické membráně, ale to už je jiný příběh… Záměna jediné aminokyseliny u bílkoviny ARF-GEF tak určuje, zdali bude citlivá k BFA či nikoli, tudíž tato změna funguje jako jakýsi „přepínač“.